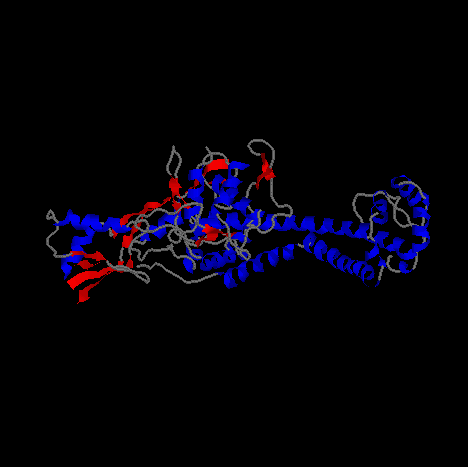
Pictured above and below is Gyrase from different angles.
The image is a cartoon representation with helical segments of the protein are
shown in blue and beta sheet regions are shown in red.
Topoisomerase
When the double helix of DNA, which is composed of two strands, separates, helicase makes these two strands rotate around each other. But there is a problem due to the topological reason that the unreplicated part ahead of the replication fork will rotate around its helical axis when the two strands separate at the replication fork (1). It causes strong strain in the helix (1). Thus, it is impossible to unlink the double helical structure of DNA without disrupting the continuity of the strands. In order to perform unraveling of a "compensating winding up" DNA, enzymes are required (1). Topoisomerase changes the linking number as well as catalyzes the interconversionn of other kinds of topological isomers of DNA (2).
- Topoisomerase I (w protein)
Type-I enzymes change the linking number by breaking a transient single-strand in duplex DNA without consumption of the energy of the phosphodiester bond (2). Type I enzymes catalyze the relaxation of closed circular, negatively supercoiled DNA. But E.coli topoisomerase I can partially remove the negative turns and binds to ssDNA and catalyzes the knotting of single-stranded circles (2). E.coli topoisomerase I does not relax positive supercoils under normal circumstances (2). E.coli topoisomerase I breaks single-stranded homopolymeric oligonucleotides of dA, dT, and a lesser extent dC at the same time (2). The enzyme attaches at +1 position and prefer a C residue 4 bases 5' to the break site, -4 position (2). According to Champoux, the E.coli topoisomerase I passes the unbroken strand through a gate that is formed when the enzyme attaches to the two ends of the DNA at the transient break and then rejoin the DNA strand (2).
- Type-II topoisomerase (E. coli gyrase)
Type II enzymes break the transient double-strand DNA so that the linking number is changed in steps of two (3). The type II DNA topoisomerases have the ability of "relaxation/supercoiling," "catenation/decatenation," and "knotting/unknotting." Bacterial gyrases are composed of heterotypic tetramers (A2B2). Subunits A (~100kD) and subunits B (~90kD) are encoded by the gyrase gyrA and gyrB genes (3, 4). Subunit B is involved in the ATP hydrolysis and energy transduction, while subunit A plays a role in the DNA breakage and reunion (3, 4). The DNA gyrase of E. coli facilitates the initiation step by its negative supercoiling of DNA that could promotes the unpairing of the strands before a helicase (4). Gyrase hydrolyzes ATP to ADP and Pi in the presence of DNA (4). This reaction is highly specific for ATP (4). Gyrase also relaxes negative supercoils in the absence of ATP but cannot relax positive supercoils (4). Mechanism of supercoiling is as follows (4). Gyrase binds to a DNA at specific sequences so that the segments form a right-handed nodes in the upper part and stabilize a positive supercoil and induce a "counterposing negative supercoil" in the lower node. The binding of ATP by the gyrB promoters brings the conformational change of gyrase that stabilizes a left-handed node. As results of the strain, gyrase breaks a dsDNA at the back of the right-handed node and passes the front segment through the gap, which change the sign of the node. Broken two ends of DNA do not rotate because gyrase holds both ends. The gyrA promoters are combined by covalent bonds with each 5' phosphoryl terminus of the break. Finally, the break is sealed by using the conserved energy from the broken phosphodiester bonds. Then, gyrase catalyzes hodrolysis of ATP and the passed segment is released for resetting. The enzyme goes back to its original conformation due to the desorption of the loosely bound ADP. When negative supercoiling density is increased, which makes binding of gyrase to DNA, the enzyme is released.
- Discovery of DNA gyrase
DNA gyrase from E. coli was discovered by Gellert and his colleagues in 1976.
According to Gellert, during the recombination of phage l DNA from E. coli, the
process requires negatively supercoiled DNA substrate (5). But if ATP is
provided sufficiently with E. coli cell fraction, relaxed closed-circular DNA
could replace the negatively supercoiled DNA substrate (5). From the results of
this experiment, they concluded that E. coli extract contains an ATP-dependent
activity that converts relaxed closed-circular DNA to the supercoiled DNA (5).
They purified the enzyme that has such ability and called it DNA gyrase (5).
Materials and methods that Gellert used in his experiment are introduced in his
article, "DNA gyrase: An enzyme that introduces superhelical turns into DNA"
(5).
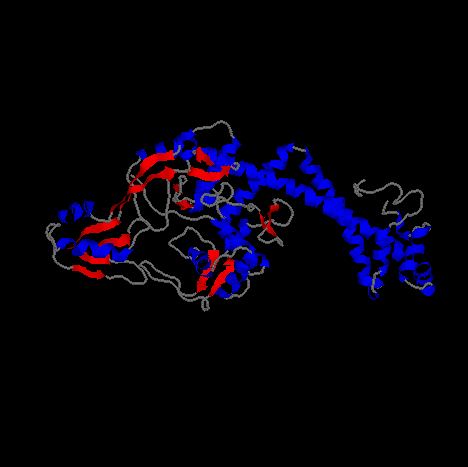
Pictured above and below is Gyrase from different angles.
The image is a cartoon representation with helical segments of the protein are
shown in blue and beta sheet regions are shown in red.
Reference:
Weaver, Robert F. Molecular biology. 1999. 650-651.
Champoux, James J. "Mechanistic Aspects of Type-I Topoisomerases." In Cozzarelli, Nicholas R. and Wang, James. C. (Ed.), DNA topology and is biological effects. pp. 217-242.
Hsieh, Tao-shih. "Mechanistic Aspects of Type-II DNA Topoisomerases." In Cozzarelli, Nicholas R. and Wang, James. C. (Ed.), DNA topology and is biological effects. pp. 243-263.
Cozzarelli, N. R. 1980. science 207:953-960.
Gellert, Martin; Mizuuchi, Kiyoshi; O'Dea, M. H.; Nash, H. A. 1976. Proc. Natl. Acad. Sci. USA. 73:3872-3876.